




キビコクNEWS
【農学部】桧原教授の研究成果が、国際科学雑誌「Science」に掲載
- 農学部
茎の節と節間ができるしくみを解明
~植物科学の未踏の地「茎の発生学」に挑む~
概要
節と節間の繰り返し構造からなる「茎」の発生メカニズムは、植物の主要器官の発生メカニズムの中で唯一研究が進んできませんでした。これは、多くの植物種で節と節間が不明瞭で、形態的特徴に乏しいことが理由として考えられます。
情報・システム研究機構 国立遺伝学研究所の津田勝利助教、前野哲輝技術専門職員、野々村賢一准教授、および広島大学の田中若奈准教授、吉備国際大学の桧原健一郎教授らの共同研究グループは、節と節間の区別が明瞭なイネの茎に着目し、茎の基本パターンが損なわれた矮性変異体を解析することで、茎の発生メカニズムの解明に挑みました。
その結果、(1)節は、「葉」の発生プログラムが「茎」に介入することで生まれること、(2)節間は、「葉」の発生プログラムの介入を「茎頂」の発生プログラムが適度に制限することで生まれること、(3)これらの発生プログラムのメカニズムは、3次元的な枝構造しか持っていなかった種子植物の祖先が葉を獲得する過程で生じた可能性が高いこと、を見出しました(図1)。
本研究は、植物発生学における最後の砦とも言える茎の基本発生プログラムとその進化過程を明らかにしました。本成果によって、農作物の収量に影響する茎形質の改良につながることが期待されます。 本研究成果は、国際科学雑誌「Science」に2024年6月14日(日本時間)に掲載されます。
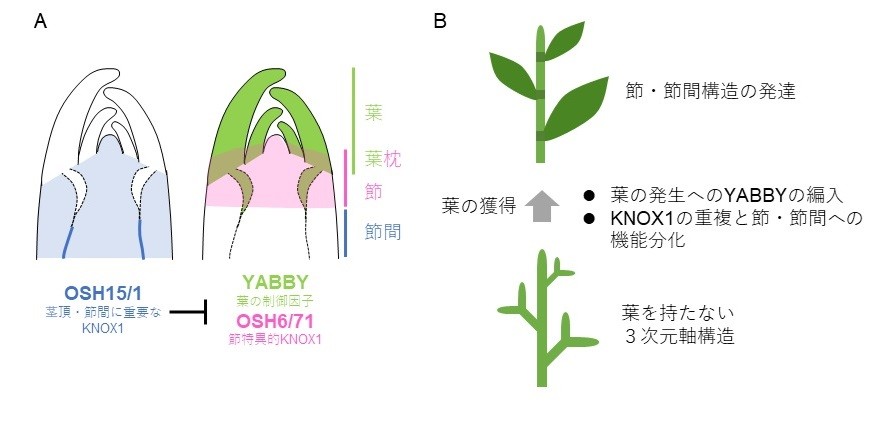
図1: イネの茎における節と節間の発生メカニズム
(A)葉の制御遺伝子「YABBY」と機能分化した「KNOX1遺伝子ファミリー」の発現領域により節と節間が規定される。
(B) 上記のメカニズムは種子植物の祖先が「葉」を獲得した際に生じ、養分交換のための節と伸長に特化した節間を区別して発達させることができるようになったと考えられる。
成果掲載誌
本研究成果は、国際科学雑誌「Science」に2024年6月14日(日本時間)に掲載されます。
論文タイトル: YABBY and diverged KNOX1 genes shape nodes and internodes in the stem.
(YABBYと機能分化したKNOX1遺伝子は茎の節と節間を形成する)
著者: Katsutoshi Tsuda*, Akiteru Maeno, Ayako Otake, Kae Kato, Wakana Tanaka, Ken-Ichiro Hibara, and Ken-Ichi Nonomura
(津田勝利*、前野哲輝、大嶽文子、加藤夏恵、田中若奈、桧原健一郎、野々村賢一) * 責任著者
研究の詳細
研究の背景
茎は植物地上部のすべての器官を繋ぎ支える軸構造で、節と節間を基本単位としています(図2A,B)。節は葉や花と茎の連絡点であり、伸長せず、複雑な維管束網を通じて精密な養水分の交換を行います。一方、節間は伸長に特化しており、効率的な光合成や花粉・種子散布に貢献します(図2C,D)。水没時に伸長する浮きイネの茎、タケの地下茎、イモの貯蔵器官といったように、茎は植物の多様な生存戦略をも担っています。
一方で、二十世紀後半に穀物の倒伏耐性と収量を劇的に増加させた「緑の革命」では、節間が短くなった矮性変異が活用されており、茎の構造は農業の発展に大きく関わってきました。
このような重要性にも関わらず、茎の発生メカニズムはほとんど研究されていません。これは主に、節と節間は多くの種で外見上の形態的特徴が乏しく、発生学的なアプローチが困難であったためと考えられます。
本研究で着目したイネでは、節と節間の区別が明瞭です(図2B)。そこで本研究グループは、イネが茎の発生メカニズム研究の良いモデルになると考え、イネを用いて節と節間が生じるメカニズムを解析しました。

図2: イネの茎の構造.
(A) イネの植物体の概略図。円柱は茎を示し、繰り返し単位ごとに色を塗り分けている。
(B) 発達中の幼い茎と葉。点線は内包される上位の葉。
(C) 茎の伸長の様子: 節間のみ伸長する。
(D) 節と節間の断面図: 節には葉と茎の間での養分交換に特化した肥大維管束(Enlarged Vascular Bundle (EVB):矢頭)が特異的に形成される。
本研究の成果
茎頂分裂組織(SAM)(1)は、地上部器官を生み出す組織であり、KNOX1遺伝子ファミリー(2)がコードする転写因子の作用により茎頂分裂組織が維持されます。KNOX1遺伝子ファミリーの一つであるOSH15遺伝子が欠損したd6変異体では節間が短くなることが知られていました。d6変異体の茎の内部構造を、CTスキャンを用いて3D構築したところ、節の特徴が広がることで、節間が短くなっていることがわかりました。さらに、OSH15遺伝子のパラログ(3)であるOSH1遺伝子の機能を部分的に欠損させる(d6 osh1/+)と、茎全体が節化し、節間が消失しました。このことから「OSH15・OSH1」サブクレード(4)は節の分化を適切な範囲にとどめ、節間形成を促すことがわかりました(図3)。

図3: d6・d6 osh1/+ 変異体では節が広がり、節間が消滅する.
(A) 若い茎のCTスキャン画像。上段は外観、下段は内部構造を示す。括弧は節の特徴であるEVBが発達した領域を示す。
(B) 茎の横断切片。矢頭: EVB、矢印:分散維管束、括弧:内皮層、を示す。ともに野生型では節のみに形成されるが、変異体ではそれ以外の部位でも異所的に形成されている。
(C) これらの結果に基づき、「OSH15・OSH1」サブクレードは節に分化する領域を制限することが示唆された。
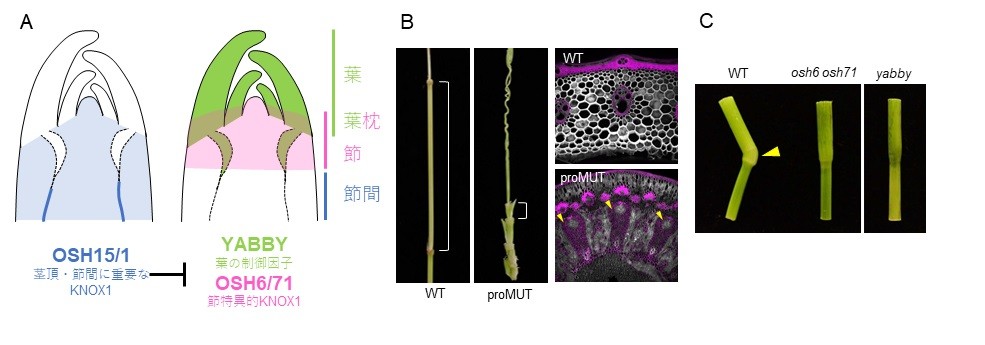
図4: YABBY遺伝子と機能分化したKNOX1遺伝子ファミリーによる節・節間形成機構
(A) YABBY遺伝子およびKNOX1遺伝子ファミリーの発現ドメインが「節(+葉枕)」と「節間」の領域を規定する。これまで葉と茎頂の制御で知られていた因子の新たな側面が明らかとなった。
(B) 遺伝子配列を変化させることにより、OSH15タンパク質がYABBY遺伝子に作用できなくなった変異体(proMUT)では、節間が節化し、本来存在しないEVB(矢頭)が出現する。この表現型は、d6 osh1/+変異体の表現型に酷似する。
(C) 節特異的KNOX1であるOSH6・OSH71変異体やYABBY変異体では、節の特徴である葉枕(矢頭)が形成されず、茎が重力屈性を発揮できなくなる。
興味深いことに、系統解析の結果、KNOX1遺伝子ファミリーのサブクレードの分化は被子植物と裸子植物の共通祖先で起こったことがわかりました。種子植物の祖先は葉を持たず、3次元的な枝構造のみからなっていたこと、またYABBYは種子植物の葉の獲得に重要な役割を果たしたことが知られています。従って、初期の種子植物でKNOX1遺伝子が重複により増え、それぞれの遺伝子が機能分化したことは、葉の獲得と並行して養分交換のための節を発達させつつ、節間伸長による空間支配を可能とするために重要であったと考えられます(図5)

図5: 節と節間の進化
種子植物は葉のない3次元軸構造から独自に葉を進化させた。その際にYABBYが葉の発生プログラムに組み込まれた。 KNOX1の節・節間への機能分化は被子植物と裸子植物の共通祖先で起こっており、初期の種子植物が葉を進化させた過程で遺伝子の重複・機能分化が起こったと考えられる。
今後の期待
本研究により、植物発生学の中で唯一取り残されていた茎の発生学の突破口が開かれました。茎頂分裂組織や葉の発生に関わるとされてきた遺伝子の新たな機能が浮き彫りになったことで、関連遺伝子の研究が進み、より詳細な植物の器官メカニズムが今後明らかになるでしょう。茎の発生機構とその多様性の理解は、さまざまな農作物における理想的な茎形質コントロールにつながることで育種に貢献することが期待されます。
用語解説
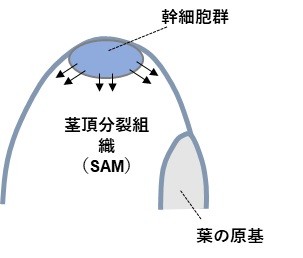
(1) 茎頂分裂組織 (Shoot Apical Meristem: SAM)
茎の先端に位置するドーム状の未分化な組織で、先端に茎頂幹細胞を有する。幹細胞から生み出された娘細胞は、新たに生じた娘細胞によりドームの側面・基部へと押し出され、ドームから切り出される形で分化が開始し、後に葉・茎・腋芽へと分化する(図5)。
(2) KNOX1遺伝子ファミリー
Class I knotted1-like homeobox (KNOX1)遺伝子ファミリーは、茎頂分裂組織の未分化状態を維持し、側生器官(葉や花など)の分化を抑制することで知られる。茎頂分裂組織と若い茎組織で発現するが、葉などが生じる予定領域では消失する。イネのKNOX1は大きくOryza sativa homeobox15 (OSH15)・OSH1のサブクレードと、OSH6・OSH71のサブクレードに分かれる。OSH6/71サブクレードの機能は未知であった。(図7)
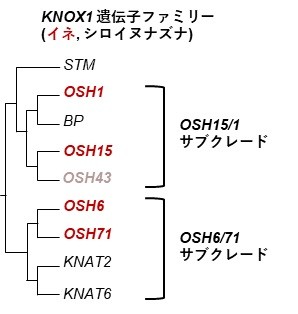
図7. Class I KNOX遺伝子ファミリー
イネとシロイヌナズナのKNOX1の系統樹. 節間形成に重要なOSH15/1サブクレードと節特異的なOSH6/71サブクレードに分かれる.(注:イネではSTMは失われている. OSH43はほとんど発現していない)
(3) パラログ
遺伝子重複によって生じた相同遺伝子。
(4) サブクレード
「種(しゅ)」と「遺伝子」いずれの場合にも「共通の祖先」から派生したすべての子孫からなる集団を「クレード」と呼び、さらにその部分集団を「サブクレード」と呼ぶ。
(5) YABBY遺伝子
葉の分化・発達に不可欠なことで知られる転写因子。種子植物では遺伝子数が増加し高度に保存されているが、その他の系統では散発的にしか見つからないことから、種子植物が葉を獲得させた際に重要な役割を果たしたと考えられている。
(6) 葉枕
植物の葉の基部に(節に隣接して)形成される構造で、光や重力に応答して屈性を発揮する。イネでは、植物体が倒れた際に重力に対して反対方向に曲がり、植物体を起き上がらせる役割を持つ。葉の一部と考えられてきたが、本研究から葉と節の制御因子が協働して葉枕が形成されることがわかった(図3C)。
研究体制と支援
本研究は、情報・システム研究機構 国立遺伝学研究所・植物細胞遺伝研究室が中心となり、広島大学・吉備国際大学との共同研究によりおこなわれました。
またこの研究は、日本学術振興会(JSPS)科研費(18H04845, 20H04891, 22H02319, 23H04754, 21H04729)の支援を受けておこなわれました。
問い合わせ先
<研究に関すること>
国立遺伝学研究所 植物細胞遺伝研究室 助教 津田勝利 (つだ かつとし)
広島大学大学院 統合生命科学研究科 植物遺伝学研究室 准教授 田中若奈 (たなか わかな)
吉備国際大学農学部 植物遺伝学研究室 教授 桧原 健一郎 (ひばら けんいちろう)
<報道担当>
国立遺伝学研究所 リサーチ・アドミニストレーター室 広報チーム
TEL: 055-981-5873
広島大学 広報室
TEL:082-424-6762 FAX:082-424-6040
学校法人順正学園 吉備国際大学 入試広報室
TEL: 086-231-3600
※Zoom会議での個別取材にも対応できますので、Zoom会議をご希望の場合には、その旨お知らせください。
植物サイズを制御する遺伝子の研究/ソルガム種子の遺伝学的研究/柑橘種子に関する研究
教授桧原 健一郎ひばら けんいちろう
- 主な担当授業科目:植物バイオテクノロジー概論、分子生物学、フィールド実習I、II
- 詳細を見る
